
科学界为什么,为谁捂盖新冠病毒来源真相(中)
.接续:科学界为什么,为谁捂盖新冠病毒来源真相?(上)
https://web.6parkbbs.com/index.php?app=forum&bbsid=2060&act=view&tid=2510400本文的评析对象是如下法广(RFI)报道:
新冠溯源: 科学家们为何要压制实验室泄密理论?
https://www.rfi.fr/cn/%E5%9B%BD%E9%99%85/20220114-%E6%96%B0%E5%86%A0%E6%BA%AF%E6%BA%90-%E7%A7%91%E5%AD%A6%E5%AE%B6%E4%BB%AC%E4%B8%BA%E4%BD%95%E8%A6%81%E5%8E%8B%E5%88%B6%E5%AE%9E%E9%AA%8C%E5%AE%A4%E6%B3%84%E5%AF%86%E7%90%86%E8%AE%BA该报道通篇“无数次”使用概念偷换、事实偷换等欺骗手法,反复阉割、篡改事实,无中生有,一而再,再而三地避重就轻、转移视线、栽赃嫁祸,以捂盖新冠病毒来源的内在真相。下面继续展示、剖析报道中的谎言、诡辩、欺诈,捂瞒、误导、欲盖弥彰。谎言五报道中说:
根据法拉尔的一份在2月2日星期日发给美国国家卫生研究院院长弗朗西斯-柯林斯和美国国家过敏和传染病研究所所长安东尼-福奇的电子邮件的记载,杜兰大学的罗伯特-加里(Robert Garry )和斯克里普斯研究所的迈克尔-法赞-的关注点集中在SARS-CoV-2基因组的一个特征上,这个特征以前从未在任何其他类似SARS的冠状病毒中出现过:插入了一个12个字母的基因序列(与蝙蝠中最接近的病毒相比),创造了一个叫做furin酶切入点使得病毒的传染性大大增强。Farzan认为,"在实验室外很难解释这个问题",Garry "想不出一个合理的自然情景......想不出这在自然界是如何完成的"。。。微生物学家Andrew Rambaut 也认为从微生物演变的角度来看,病毒酶切入点的出现是“十分不寻常的”!并且暗示只有武汉病毒学者才能够作出解释。“微生物学家Andrew Rambaut 。。。暗示只有武汉病毒学者才能够作出解释”这句话,要么是Andrew Rambaut罔顾事实、掩耳盗铃的恶毒栽赃,要么是RFI报道无中生有、凭空杜撰的下作诬蔑。能解释新冠病毒中furin酶切位点的人,应该是做过furin酶切位点研究的人。武汉病毒研究所应该为新冠的furin酶切位点作出解释吗?武汉病毒研究所从未做过在冠状病毒中插入furin酶切位点的研究、实验,你叫他们怎么解释?真正能解释,真正应该解释新冠病毒中furin酶切位点来源的人,不是武汉病毒学者,而是美国病毒学家们。因为美国病毒学家们十多年来做过不计其数的在冠状病毒或其它病毒中插入furin酶切位点的病毒改造研究、实验,美国病毒学家对furin酶切位点的研究、实验极为深入、全面,涵盖以下几种情况:
1、在没有furin酶切位点的(冠状)病毒中插入一个或多个furin酶切位点;
2、在已有furin酶切位点的(冠状)病毒中插入新的furin酶切位点;
3、改造(冠状)病毒中已有的furin酶切位点的酶切组合,对比改造前后不同furin酶切组合的水解、切割效率,致病能力、效果。。。举两个例子。第一个例子。2006 年3月,Montana大学的Jack Nunberg等人在Elsevier上发表了如下论文:
Furin cleavage of the SARS coronavirus spike glycoprotein enhances cell–cell fusion but does not affect virion entry
SARS病毒刺突(糖)蛋白的弗林切割增强了细胞-细胞融合,但不影响病毒进入
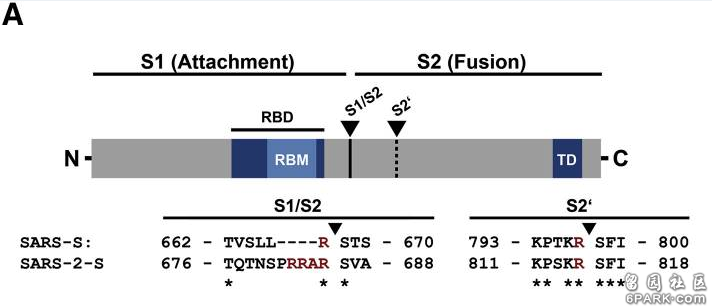
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7111780/该研究在SARS-CoV刺突蛋白(Spike蛋白,S蛋白)S1/S2位置处(S蛋白S1、S2亚基交界处)插入了一个furin(弗林)酶切位点,并实验证明,这一FCS(furin cleavage site,即furin酶切位点)插入显著增强了SARS病毒诱导宿主细胞发生细胞–细胞融合,形成多核扩大细胞--合胞体的能力(细胞-细胞融合形成合胞体能提高病毒在宿主组织、器官中的扩散效率)。新冠病毒的furin酶切位点也位于S蛋白S1、S2亚基交界处,与该论文的furin酶切位点插入位置完全相同。
新冠病毒(SARS-2)S1/S2位置处的“RRAR”furin酶切位点
SARS病毒本身没有S1/S2处的furin酶切位点,
与新冠同谱系、同支系(冠状病毒β谱系B支系)的新冠近亲病毒全都没有S1/S2处的furin酶切位点;
与新冠同谱系(冠状病毒β谱系)的蝙蝠冠状病毒全都没有S1/S2处的furin酶切位点;
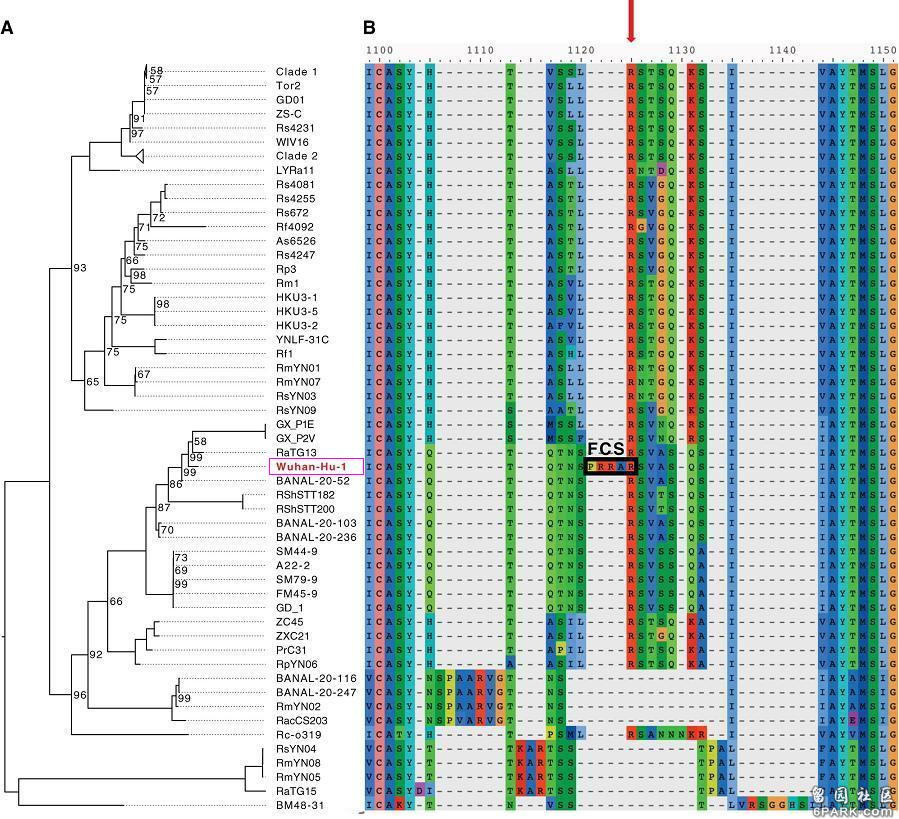
与新冠刺突蛋白相似度>40%的新冠所有天然近亲、远亲也全都没有S1/S2处的furin酶切位点。上图是新冠病毒(Wuhan-Hu-1毒株)与进化树中的“亲戚”冠状病毒在S1/S2附近氨基酸序列的对照图,最上方的红色箭头指示的就是S1/S2位置(S1、S2亚基交界处)。由图可见,在S1/S2处有furin酶切位点的唯有新冠病毒,图中的新冠近亲、远亲,无一在在S1/S2处有furin酶切位点(全都缺失furin酶切位点对应的氨基酸)。请大家作个判断,新冠病毒S1/S2处的furin酶切位点是怎么得来的?是自然演化产生的,还是人为插入的?第二个例子。2009年4月,Cornell大学的加里·惠特克(Gary Whittaker)等人在PNAS(美国国家科学院院刊)上发表了如下论文:
Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites
经由两个不同位点的连续蛋白水解切割(触发)的SARS 病毒刺突蛋白激活(活性增强)
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2660061/该研究在SARS病毒刺突蛋白的两个位置插入了furin酶切位点,一个在S1/S2处,另一个在S2亚基内(SARS刺突蛋白的第793~797氨基酸之间),并构造了三个SARS病毒的人工变异(改造)体,其中两个各含一个furin酶切位点插入,另一个兼具两个furin酶切位点插入。研究实验证明:
1、S1/S2位置插入的furin酶切位点可使S蛋白被有效水解切割,导致S1、S2亚基相互分离;
2、S1/S2位置插入的furin酶切位点显著增强了细胞-细胞融合,促进了合胞体的生成;
3、S1/S2位置插入的furin酶切位点显著提高了SARS病毒的感染效率;
4、S2′位置插入的furin酶切位点可进一步增强细胞-细胞融合,促进合胞体生成,兼具两个furin酶切位点的SARS改造体可导致超过95%的(被感染组织中的)细胞发生细胞-细胞融合,形成非常大的合胞体。美国科学家所做的furin酶切位点相关的研究、实验还有很多,我阅读过的研究例证,将汇集、整理在稍后的其它文章中。对新冠病毒S1/S2位置处,其近亲无一具备的furin酶切位点,谁能够作出解释,谁应该作出解释,作出澄清?是报道所说的“武汉病毒学者”,还是做过大量furin酶切位点插入、改造实验的美国病毒学家们?谎言六报道中说:
从最新公布的邮件上可以看出,美国国家过敏和传染病研究所所长安东尼-福奇从第一时间便警告专家不应该公开讨论病毒来自实验室的理论,因为这会对科学研究,尤其是中国的科学研究造成攻击。这也是为何福奇本人曾经十多次强调病毒只能是自然来源,而美国国家卫生研究院院长弗朗西斯-柯林斯更是将实验室泄露理论定性为荒谬。“自然来源论”的反面是“实验室来源论”,不是“实验室泄漏论”。由艾滋病、西班牙流感的先例可知,病毒的最早发现地、最早暴露地不等于病毒的来源地、产生地(设计、制造地)。如果新冠病毒不是自然产生的,那么,它就是实验室产生的;如果新冠病毒是实验室产生的,那么,它可能来自武汉的某个实验室,它同样可能来自武汉之外、中国之外的某个实验室。没有任何证据,没有任何理由可排除新冠病毒来自武汉之外实验室的可能性。新冠病毒来自武汉之外实验室的可能性,是新冠溯源工作应深入调查,不可忽略、不容遗漏的必要内容。
“实验室泄漏论”只是“实验室来源论”的一个分支。来自实验室,不等于来自“实验室泄漏”。“实验室泄漏”,相当于“本地实验室泄漏”,“实验室泄漏论”的潜台词是:新冠病毒来自武汉的某座实验室。用“实验室泄漏论”偷换“实验室来源论”,将二者混为一谈,是将“实验室来源论”蕴含的多种可能性阉割为“武汉实验室来源”单一可能性,是在隐蔽、捂盖新冠病毒的其它实验室来源可能性。在以往的文章中,我曾从不同角度,以不同方式,基于不同证据,多次论证过以下两个结论(最近的一次论证是在上篇文章的“谎言一”小节给出的):
1、新冠病毒不是武汉病毒研究所设计、制造的;
2、新冠病毒不可能是武汉病毒研究所泄漏的;这是两个得到大量坚实证据交叉支撑,经得起反复推敲的结论。由这两个结论可知,“实验室泄漏论”根本不成立!美国及美国的盟友们为什么无休止地炒作“实验室泄漏论”?他们为什么对否定“实验室泄漏论”的众多铁证视而不见?为什么美国政府一直抱怨缺乏足够的溯源证据、资料?因为美国溯源调查的目的不是揭示、还原新冠病毒来源真相,新冠疫情产生真相;美国需要的是制造替罪羊的证据,不会引火烧身的证据,所有不利于美国,不利于美国制造替罪羊,不利于美国甩锅嫁祸的证据,统统不是证据。新冠病毒来自实验室,它又不是武汉病毒研究所设计、制造、泄漏的。那么,新冠病毒肯定来自武汉病毒研究所之外的实验室,它极有可能来自武汉之外、中国之外的实验室!弗朗西斯-柯林斯(Francis Collins)力挺的是“自然来源论”,他曾在2020年3月26日发表名为《基因研究显示 新冠肺炎病毒起源于自然》的博客文章,宣称:该病毒是自然产生的。
美国国立卫生研究院院长柯林斯:新冠肺炎病毒源于自然_新闻中心_中国网
http://news.china.com.cn/2020-03/28/content_75871483.htm对弗朗西斯-柯林斯来说,正论是“自然来源论”,谬论是“实验室来源论”。RFI报道再一次用“实验室来源论”的分支“实验室泄漏论”以偏盖全、偷换“实验室来源论”,将新冠病毒的来源与武汉病毒研究所单一挂钩,籍此隐匿、掩藏新冠病毒的其它实验室来源可能性。压制“实验室泄漏论”,是为武汉病毒研究所打掩护,是为中国打掩护;压制“实验室来源论”,是为从事冠状病毒功能增益改造研究的实验室打掩护,是为开展举国性质危险病原体功能增益改造研究,以联邦资金资助本国功能增益改造研究的国家打掩护。弗朗西斯-柯林斯、安东尼-福奇全力压制的,是“实验室泄漏论”,还是“实验室来源论”呢?他们是在为武汉病毒研究所、为中国打掩护,还是在为美国实验室,为美国打掩护呢?报道中说,安东尼-福奇第一时间警告专家不应该公开讨论病毒来自实验室的理论,福奇本人曾经十多次强调病毒只能是自然来源,美国国家卫生研究院院长弗朗西斯-柯林斯更是将实验室泄露理论定性为荒谬。二人如此积极、如此投入,如此不遗余力地压制、捂盖、撒谎,不惜身败名裂也要欺骗全世界,他们真的是为保护中国的科学研究吗?他们真的是为武汉病毒研究所打掩护吗?他们可能为中国如此捂盖新冠病毒的来源真相吗?这合乎情理吗?这说得通吗?他们是忘我奉献、舍已为人的“活雷锋”,为中国两肋插刀、至死不渝的“白求恩”吗?谎言七报道中说:
2020年3月17日自然杂志发表的标题为“The proximal origin of SARS-CoV-2”的文章对病毒来源问题一锤定音,文章虽然也对病毒基因序列组成与furin酶切入点提出疑问,但认为可以在自然演变中完成。
实验室泄露的可能性被彻底排除在外,文章的签名者中就包括微生物学家Andrew Rambaut 与病毒学家罗伯特-加里(Robert Garry )等对病毒持有极大怀疑的专家,究竟是何等原因使他们在一个月内改变看法呢?罗伯特-加里日前向美国拦截组织The Intercept.表示他当初对酶切入点的印象使(是)错误的,他说科学就是如此逐渐走向进步的,无人故意要愚弄公众,他今天依然认为他们在自然杂志上发表的文章是有关新冠病毒的最完美的分析文章。
(报道引用结束)Nature论文“The proximal origin of SARS-CoV-2(SARS-CoV-2的近端起源)”地址如下:
https://www.nature.com/articles/s41591-020-0820-9中文介绍可参考:
Nat Med:研究证实SARS-CoV-2属自然进化,不可能是人为制造! - 组学专区 – 生物谷
https://news.bioon.com/article/6752206.html美英澳科学家《自然医学》再添力证:新冠病毒乃自然进化产物…… 广东省科学技术厅
http://gdstc.gd.gov.cn/kjfyzl/gnwdt/content/post_2936176.html论文力撑自然来源论,宣称:对SARS-CoV-2及相关病毒的基因组序列进行分析后发现,没有证据表明该病毒是实验室制造或实验室(方式)产生的。显而易见,论文否定、排除的不是“实验室泄露论”,而是“实验室来源论”。RFI报道第N次使用概念偷换、事实偷换手法,以偏代全地谎称“实验室泄露的可能性被彻底排除在外”。这么一偷换,一篇捂盖新冠“实验室来源”,为美国危险病原体功能增益改造研究打掩护的论文,摇身一变,变成了捂盖“武汉实验室泄漏”,为武汉病毒研究所打掩护的论文;论文的包庇、服务对象,由开展功能增益研究的美国实验室,变成了没有做过功能增益研究,没有发表过任何功能增益研究论文的武汉病毒研究所;论文的包庇、服务国,从开展举国性质功能增益研究的第一生物科技强国--美国,变成了功能增益研究屈指可数、生物科技水平落后美国十几年的中国。经偷换、篡改捏造出的包庇、服务对象作为新冠病毒来源的唯一嫌疑犯被RFI押上了被告席,而论文真正的包庇、服务对象则悄然隐身于世人目光之外。论文的作者是以下五人:美国斯克利普斯研究所的克里斯蒂安·安德森(Kristian G Andersen)、英国爱丁堡大学进化生物学研究所的Andrew Rambaut、美国哥伦比亚大学公共卫生学院感染与免疫中心主任W.伊恩·利普金(W. Ian Lipkin)、澳大利亚悉尼大学的爱德华·霍姆斯(Edward C. Holmes),美国杜兰大学医学院微生物学和免疫学系的罗伯特·f·加里(Robert F. Garry)。五人中三人是美国病毒学家,另两人分属英国、澳大利亚。“巧合”的是,五人的所属国澳大利亚(AU)、英国(UK)、美国(US)恰好是AUKUS军事联盟(澳英美军事联盟)的三个成员国。AUKUS军事联盟的首要战略敌人是中国,美、澳、英三国是近几年打击中国现政府最不遗余力的三个国家。试想,这三个国家的病毒学家可能联手为武汉病毒研究所打掩护,联手为中国捂盖新冠病毒的来源真相吗?他们究竟在为谁捂盖?五位论文作者中,当初认为新冠病毒来自实验室的至少有三位。上述报道文字提到了两位:爱丁堡大学的Andrew-Rambaut,杜兰大学的罗伯特·加里(Robert Garry)。还有一位是上篇文章提到过的,曾在2020年1月31日致信安东尼-福奇,表示新冠病毒有基因改造特征,不符合进化理论,但在次日与福奇电话会议后迅速改变立场的斯克利普斯研究所的克里斯蒂安·安德森(Kristian G Andersen)。甘当政客们的科学工具,可以随时出卖真相、事实、是非的三个墙头草有科学操守和基本人格可言吗?报道中提到,罗伯特-加里说自己并非“故意要愚弄公众”,并声称“他今天依然认为他们在自然杂志上发表的文章是有关新冠病毒的最完美的分析文章。”如此“完美”的论文,不近距离欣赏、领略一番将是巨大的遗憾。罗伯特-加里等五人的Natue大作到底有多完美呢?它至少有三大“完美表现”。完美表现一论文称,新冠病毒的骨架与已知病毒的骨架都有很大不同,因此,它不是基于某种已知病毒骨架构建的(进而它不是实验室构建的)。论文隐含了一个假设:实验室设计、制造不出与已知病毒的骨架都有很大不同的病毒。这一假设的推论是:与已知病毒的骨架都有很大不同的病毒一定不是实验室构造的。论文使用了上述假设的推论来否定新冠病毒来自实验室。问题是,上述假设是错误的。实验室能不能设计、制造出与已知病毒的骨架都有很大不同的病毒呢?太能了!对现代基因改造技术来说,这根本就不是个问题。我举个例子。2008年12月16日,Ralph S. Baric(冠状病毒研究全球第一人,冠状病毒功能增益改造狂人)领导的团队发表了一篇PNAS(美国国家科学院院刊)论文:
Synthetic recombinant bat SARS-like coronavirus is infectious in cultured cells and in mice
合成的重组蝙蝠类SARS冠状病毒在培养细胞和小鼠中具有传染性。
https://www.pnas.org/content/105/50/19944论文通过两步改造,合成了一个重组病毒Bat-SRBD。Bat-SRBD是用5种病毒原材料重组合成的,它们是SARS-CoV和4种蝙蝠冠状病毒:HKU3-1、HKU3-2、HKU3-3和RP3 。重组病毒Bat-SRBD的RBD来自SARS病毒,骨架(其它部分)来自4种蝙蝠冠状病毒。实验证明,Bat-SRBD能有效感染HAE细胞(人气道上皮细胞)中的纤毛细胞(但不能感染同属HAE细胞的非纤毛细胞)、非洲绿猴的Vero E6细胞(一种肾细胞)、表达人类ACE2的转基因小鼠DBT细胞(延迟脑肿瘤细胞)DBT-hACE2、表达果子狸ACE2的转基因小鼠DBT细胞DBT-cACE2,并在以上各细胞组织中大量复制。也就是说,重组病毒Bat-SRBD是一种能感染人类、非洲绿猴、果子狸(civet)三种动物,复制能力强大,可跨物种感染、传播的实验室改造病毒。由这篇论文可知,改造病毒不必只基于一种病毒(骨架),可以基于多种病毒来获得改造病毒,改造病毒将兼具多种病毒的结构,改造病毒的结构将与素材病毒中的任何一种都有明显差异。早在2008年,病毒学家就可以很容易地设计、制造出混成式的冠状病毒了。而且,Ralph S. Baric当时就有能力设计、制造出比Bat-SRBD复杂100倍的病毒。2000年发明基于基因序列合成冠状病毒的反向遗传平台之后,Ralph S. Baric等人已将冠状病毒的设计、改造简化为基因序列文本文件的设计、改造了。因此,论文的假设是错误的,论文作者是在故意装糊涂。为了颠倒黑白,“论证”自然来源论,装糊涂是必须的。新冠病毒的骨架与已知病毒的骨架都有很大不同,原因与上述重组病毒Bat-SRBD类似。我之前的文章多次指出,新冠病毒是跨科属的病毒集大成者,它借鉴、引入、集成了多种冠状病毒、多种非冠状病毒的结构和致病特性,它不是一个基于单一原材料骨架的简单改造体。存在多重结构来源(结构来源五花八门),部分结构甚至借鉴、来自非冠状病毒(包括艾滋病病毒、埃博拉病毒等等)的新冠病毒的骨架当然与已知病毒的骨架都有很大不同。实际上,“新冠病毒的骨架与已知病毒的骨架都有很大不同”,这一事实不是新冠病毒自然产生的证据,而是新冠病毒实验室产生的证据。因为自然演化产生的病毒必然存在高度相像的祖先病毒、近亲病毒,只有在实验室经过多重改造(多重集成、多重嵌合、多重编辑)得到的病毒才会和所有自然病毒无一高度相像。完美表现二论文称,新冠病毒的RBD(Receptor Binding Domain,受体结合域,决定S蛋白即刺突蛋白ACE2结合能力的S1亚基的一部分)不是最优的,如果新冠病毒是人为设计的,那么,它的RBD应该设计得更好。论文此处的逻辑(又一个隐含的假设)非常“完美”、“严谨”:人为设计的病毒必然是最优的;不是最优设计的病毒一定不是人为设计的。按照这一逻辑,可得出如下推论:实验室设计、制造的病毒,只有终极版本才(可能)是实验室设计、制造的,其它版本都肯定不是实验室设计、制造的,肯定都是自然产生的。因为非终极版本的设计都不是最优的。新冠病毒的RBD是不是真的不够好呢?事实上,新冠的RBD设计即使不是最优的,也是极为接近最优的,这体现在:
1、新冠刺突蛋白与人类ACE2的结合亲和力(结合亲和力由刺突蛋白的RBD决定),是SARS刺突蛋白与人类ACE2结合亲和力的10~20倍;
2、新冠刺突蛋白与人类ACE2的结合亲和力,强于它与常见动物ACE2的结合亲和力,人类ACE2是新冠刺突蛋白的最佳结合对象,人体细胞是新冠病毒的最佳感染对象;
3、新冠病毒一经出现,未经过任何宿主跳跃所需的突变,其刺突蛋白便极为适合结合人类ACE2,新冠病毒便极为适合感染人类。
(由2、3可得出如下推论:新冠病毒的原始宿主很可能就是人类,新冠病毒很可能是专为感染人类而设计的。)第1点的依据是,2020年3月13日,美国德克萨斯大学奥斯汀分校Jason S McLellan团队,与美国国家过敏和传染病研究所的研究人员在《Science》杂志上联合发表的如下论文:
Cryo-EM Structure of the 2019-nCoV Spike in the Prefusion Conformation
(膜融合前构象中2019-nCoV 刺突蛋白的冷冻电镜结构)
https://science.sciencemag.org/content/367/6483/1260第2、3两点的依据是,澳大利亚弗林德斯大学、拉筹伯大学等机构的科学家2021年6月24日发表的如下Nature杂志论文(该论文早在2020年5月13日就已发表于预印本平台arxiv):
In silico comparison of SARS-CoV-2 spike protein-ACE2 binding affinities across species and implications for viral origin
(新冠刺突蛋白与ACE2结合亲和力的跨物种模拟比较及其对病毒起源的指示)
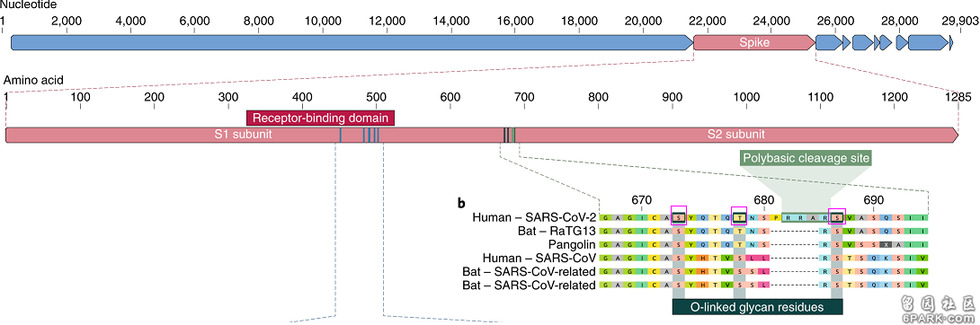
https://www.nature.com/articles/s41598-021-92388-5完美表现三论文指出,新冠病毒S1、S2亚基交界处furin酶切位点周围存在一个具有免疫屏蔽功能(可屏蔽宿主免疫系统搜索、发现、攻击)的O-Linked聚糖结构。论文称,在实验室环境中将祖本病毒置于培养细胞中连续传代不太可能变异产生这一结构,这一具有免疫屏蔽功能的结构需要在(在人体活体)免疫系统的参与下才能变异产生。因此,新冠病毒不是实验室培养产生的。论文作者又装糊涂了,他们使用了第三个不成立的假设,把实验室培养(以一定方式、压力培养病毒,促使其在连续传代过程中发生变异)作为获得实验室病毒,及人为产生O-Linked聚糖结构的唯一途径。事实上,这种正向遗传学方式早就不是病毒改造的主流手段了(有时仍作为辅助手段),它早已被直接编辑基因组或编辑基因序列的反向遗传技术取而代之了;病毒学家们根本不会,根本无须用这种难以达到既定目的的方式去产生O-Linked聚糖结构,他们可以快捷地、轻而易举地在病毒中直接构造出O-Linked聚糖结构。我们近距离观察一下新冠病毒刺突蛋白(S蛋白)中的O-Linked聚糖结构,这一结构由furin酶切位点(RRAR)两侧的三个氨基酸构成:S蛋白氨基酸序列的第673氨基酸-丝氨酸(S),第678氨基酸-苏氨酸(T),第687氨基酸-丝氨酸(S)。这三个氨基酸在下图中用紫色方框标出:
新冠furin酶切位点周围的一个苏氨酸(T),二个丝氨酸(S)形成一个O-Linked聚糖结构
新冠刺突蛋白的第678氨基酸是苏氨酸(T),而SARS和绝大多数类SARS冠状病毒(新冠和SARS同处β谱系B支系,这一支系的病毒统称类SARS冠状病毒)对应位置处的氨基酸是丝氨酸(S);新冠刺突蛋白的第673、687氨基酸则和SARS等近亲相同,都是丝氨酸(S)。
注:上图中另二个新冠近亲Bat-RaTG13和Pangalin(代表一种马来穿山甲冠状病毒)的第678氨基酸也是苏氨酸(T),与新冠相同,但这两个病毒没有病毒毒株,而且它们的基因序列是疫情发生后才宣布的,是专门用来证明新冠“自然来源论”的,它们的真实性非常可疑。基于新冠的近亲病毒(如SARS或蝙蝠冠状病毒)构造O-Linked聚糖结构,只要把对应新冠病毒刺突蛋白第678氨基酸的近亲病毒氨基酸由丝氨酸(S)替换为苏氨酸(T)就可以了(忽略O-Linked聚糖结构与furin酶切位点的关联性)。至少有两种反向遗传技术可快速实现这一替换:
一是使用基因剪刀(CRISPR-Cas9 )直接编辑近亲病毒基因组,完成氨基酸替换;
二是先编辑近亲病毒的基因序列,在基因序列(文本)中设计出这一结构(把丝氨酸的核苷酸密码子符号替换成苏氨酸的密码子符号就ok了),再基于设计好的新的基因序列使用反向遗传平台合成病毒基因组、收获病毒毒株,O-Linked聚糖结构就在所收获的病毒毒株中存在了。HIV-1病毒(即艾滋病病毒,属于逆转录病毒科、慢病毒属)、埃博拉病毒(丝状病毒科)、丙型肝炎病毒(黄病毒科)等非冠状病毒也有具有免疫屏蔽功能的O-Linked聚糖结构,新冠的O-Linked聚糖结构很可能借鉴自这些非冠状病毒。五位病毒大腕的Nature论文不是学术作品,不是写给同行看的。它是某国领导下的全球科学界的“大外宣”工具,它的存在价值是拉大旗作虎皮,作为“权威著作”供“自然来源论”反复引用, 为“自然来源论”撑腰、壮胆、开道。(未完待续)贴主:苦难与荣耀于2022_01_20 12:56:04编辑贴主:苦难与荣耀于2022_01_20 16:33:39编辑
贴主:苦难与荣耀于2022_01_21 19:48:11编辑
喜欢苦难与荣耀朋友的这个贴子的话, 请点这里投票,“赞”助支持!
请点这里投票,“赞”助支持!

已标注为苦难与荣耀的原创内容,若需转载授权请联系网友本人。若违规侵权,请联系我们
所有跟帖: ( 主贴楼主有权删除不文明回复,拉黑不受欢迎的用户 )
aroha007 给 苦难与荣耀 点“赞”支持3银元奖励!! (无内容) - aroha007 (0 bytes) 01/23/22
(^-^) 苦难与荣耀 给 aroha007 打来一扎啤酒! - 苦难与荣耀 (88 bytes) 01/25/22
感谢您大力支持! (无内容) - 苦难与荣耀 (0 bytes) 01/25/22
fuasy 给 苦难与荣耀 点“赞”支持3银元奖励!! (无内容) - fuasy (0 bytes) 01/23/22
感谢您的大力支持! (无内容) - 苦难与荣耀 (0 bytes) 01/23/22
(^-^) nuassembly 给 苦难与荣耀 赠送一尾锦鲤! - nuassembly (89 bytes) 01/22/22
(^-^) 苦难与荣耀 给 nuassembly 打来一扎啤酒! - 苦难与荣耀 (88 bytes) 01/23/22
感谢馈赠! (无内容) - 苦难与荣耀 (0 bytes) 01/23/22
(^-^) 谈今论今 给 苦难与荣耀 献上一个金话筒! - 谈今论今 (127 bytes) 01/22/22
谈今论今 给 苦难与荣耀 点“赞”支持3银元奖励!! (无内容) - 谈今论今 (0 bytes) 01/22/22
(^-^) 苦难与荣耀 给 谈今论今 赠送一瓶红葡萄酒! - 苦难与荣耀 (127 bytes) 01/23/22
感谢谈今兄! (无内容) - 苦难与荣耀 (0 bytes) 01/23/22
nicoledeng 给 苦难与荣耀 点“赞”支持3银元奖励!! (无内容) - nicoledeng (0 bytes) 01/22/22
(^-^) 苦难与荣耀 给 nicoledeng 打来一扎啤酒! - 苦难与荣耀 (88 bytes) 01/23/22
感谢您支持、鼓励! (无内容) - 苦难与荣耀 (0 bytes) 01/23/22
(^-^) 東方聖人 给 苦难与荣耀 颁发一张学霸证书! - 東方聖人 (89 bytes) 01/21/22
(^-^) 苦难与荣耀 给 東方聖人 打来一扎啤酒! - 苦难与荣耀 (88 bytes) 01/21/22
感谢东方兄馈赠! (无内容) - 苦难与荣耀 (0 bytes) 01/21/22
应该的!因为我们的坚持,让西方舆论不可以只手遮天。 (无内容) - 東方聖人 (0 bytes) 01/21/22
(^-^) 苦难与荣耀 给 東方聖人 泡上一杯枸杞菊花! - 苦难与荣耀 (89 bytes) 01/23/22
资深专家分析水平!希望能总结一下,让非专业读者能一目了然 (无内容) - august (0 bytes) 01/21/22
感谢您的赞誉! - 苦难与荣耀 (207 bytes) 01/21/22
?二不换不是说:我親自指挥親自部署的 (无内容) - 真话难听 (0 bytes) 01/21/22
逻辑性如此强大的文章,没有应该专业水平是写不出来的。 (无内容) - 铗归来 (0 bytes) 01/21/22
(^-^) 苦难与荣耀 给 铗归来 打来一扎啤酒! - 苦难与荣耀 (88 bytes) 01/21/22
?二不换不是说:我親自指挥親自部署的 (无内容) - 真话难听 (0 bytes) 01/21/22
病毒来自武毒所但病毒技术来自美国,福气掩盖事实怕引火烧身 (无内容) - pickle (0 bytes) 01/21/22
连柳叶刀都帮着摇旗呐喊, 感觉还是很遗憾的 (无内容) - pickle (0 bytes) 01/21/22
谢谢您鼎力大作,已转账500银元支持! (无内容) - fuasy (0 bytes) 01/20/22
非常感激您的大力支持! - 苦难与荣耀 (111 bytes) 01/21/22
我在枫林中哭泣 给 苦难与荣耀 点“赞”支持3银元奖励!! (无内容) - 我在枫林中哭泣 (0 bytes) 01/20/22
(^-^) 苦难与荣耀 给 我在枫林中哭泣 泡上一碗茉莉花茶! - 苦难与荣耀 (89 bytes) 01/21/22
感谢枫林版主支持、鼓励! (无内容) - 苦难与荣耀 (0 bytes) 01/21/22
谢谢原创首发,已转账200银元奖励! (无内容) - BLUESKYUK (0 bytes) 01/20/22
(^-^) BLUESKYUK 给 苦难与荣耀 献上一支玫瑰花! - BLUESKYUK (88 bytes) 01/20/22
(^-^) BLUESKYUK 给 苦难与荣耀 赠送一尾锦鲤! - BLUESKYUK (89 bytes) 01/20/22
(^-^) BLUESKYUK 给 苦难与荣耀 赠送一只吉祥小虎! - BLUESKYUK (89 bytes) 01/20/22
(^-^) BLUESKYUK 给 苦难与荣耀 送上祝福财源滚滚! - BLUESKYUK (89 bytes) 01/20/22
(^-^) BLUESKYUK 给 苦难与荣耀 送上祝福柿柿如意! - BLUESKYUK (89 bytes) 01/20/22
BLUESKYUK 给 苦难与荣耀 点“赞”支持3银元奖励!! (无内容) - BLUESKYUK (0 bytes) 01/20/22
(^-^) 苦难与荣耀 给 BLUESKYUK 送上一罐桂花米酒! - 苦难与荣耀 (128 bytes) 01/21/22
感谢版主支持、厚赠! (无内容) - 苦难与荣耀 (0 bytes) 01/21/22
打开微信,扫一扫[Scan QR Code]
进入内容页点击屏幕右上分享按钮
进入内容页点击屏幕右上分享按钮
楼主前期社区热帖:
- 再次求助!原创帖极速从首页“网络社区原创”中消失! 03/04/24
- 求助!原创文章很快从“网络社区原创”中消失 02/04/24
- 武汉病毒研究所不是新冠病毒的来源地 05/10/23
- 新冠溯源,美国之音再秀谎言报道(上) 08/09/22
- 谁设计、制造了新冠病毒(三 下) 06/28/22
- 拉尔夫·巴里克对RBD关键氨基酸的研究 06/23/22
- 谁设计、制造了新冠病毒(三 上) 06/19/22
- 隐秘红色代理人蒋介石夫人宋美龄 上 05/08/22
- 为什么有人巴不得新冠病毒在中国失控扩散 04/14/22
- 新冠人造再添铁证,“有据核查”欲盖弥彰 03/25/22
- 谁设计、制造了新冠病毒(二 下) 03/06/22
- 谁设计、制造了新冠病毒(二 中)续 02/27/22
- 谁设计、制造了新冠病毒(二 中) 02/23/22
- 科学界为什么,为谁捂盖新冠病毒来源真相(下) 01/23/22
- 科学界为什么,为谁捂盖新冠病毒来源真相?(上) 01/15/22
- 新冠furin酶切位点探源,病毒学界的集体捂瞒 01/12/22
- 谁设计、制造了新冠病毒(二 上) 01/06/22
- (迎新)谁是全球新冠虚假信息攻势的领头羊? 01/01/22
- (迎新)美国之音年终谎言报道(二) 12/29/21
>>>>查看更多楼主社区动态...