
突触动力学如何启发对大脑神经网络的认识?
导语
神经网络是复杂的生物系统,其行为受到突触可塑性的影响。突触可以分为电突触(Electric Synapse)和化学突触(Chemical Synapse)。突触可塑性是指神经元之间突触连接强度的可变性,对于学习、记忆和神经功能的塑造至关重要。在集智俱乐部「计算神经科学」读书会,香港城市大学神经科学系助理教授冯志聪(Alan Fung)介绍了突触动力学,短期突触可塑性如何调节长期突触可塑性,以及关于突触竞争的研究。本文是社区成员对此次读书会分享的整理。
研究领域:计算神经科学,神经网络,突触动力学,突触可塑性,突触竞争,连续吸引子神经网络
冯志聪| 讲者
刘泽权| 整理
这次的分享是关于突触动力学对神经网络的启发。需要了解长期突触可塑性,才能了解记忆的形成,但不能一直依赖长期可塑性,需要一个调节机制,而短期可塑性是个方案。短期突触可塑性可能调节长期突触可塑性。此次分享还介绍了作者最近的一篇关于突触竞争的文章,以及怎么在机器学习中实现类似的机制。
大脑电流的产生机制
大脑如何产生电流?化学突触是两个神经元之间的连接,其中信息通过释放和接收被称为神经递质的化学物质来传递。神经递质可以与相应类型的受体结合,而这些受体包含离子通道。神经递质的结合可导致离子通道的打开,允许离子电流进出细胞,继而改变细胞的膜电位。突触还分为突触前端和突触后端,动作电位传递过来的时候,神经递质就会释放到受体,从而打开离子通道,造成去极化(depolarization),即把电位提高,或者是超极化(hyperpolarization),即把电位降低。
神经递质里,谷氨酸(glutamate)是兴奋性的,对应受体是AMPA和NMDA,可以引起钠离子进入细胞引起去极化。另外一个主要的神经递质是GABA受体,释放它的细胞通常称为抑制性细胞,因为它会打开抑制性受体让氯离子进入细胞,把细胞的电位压得更低,让它比较难去激发。
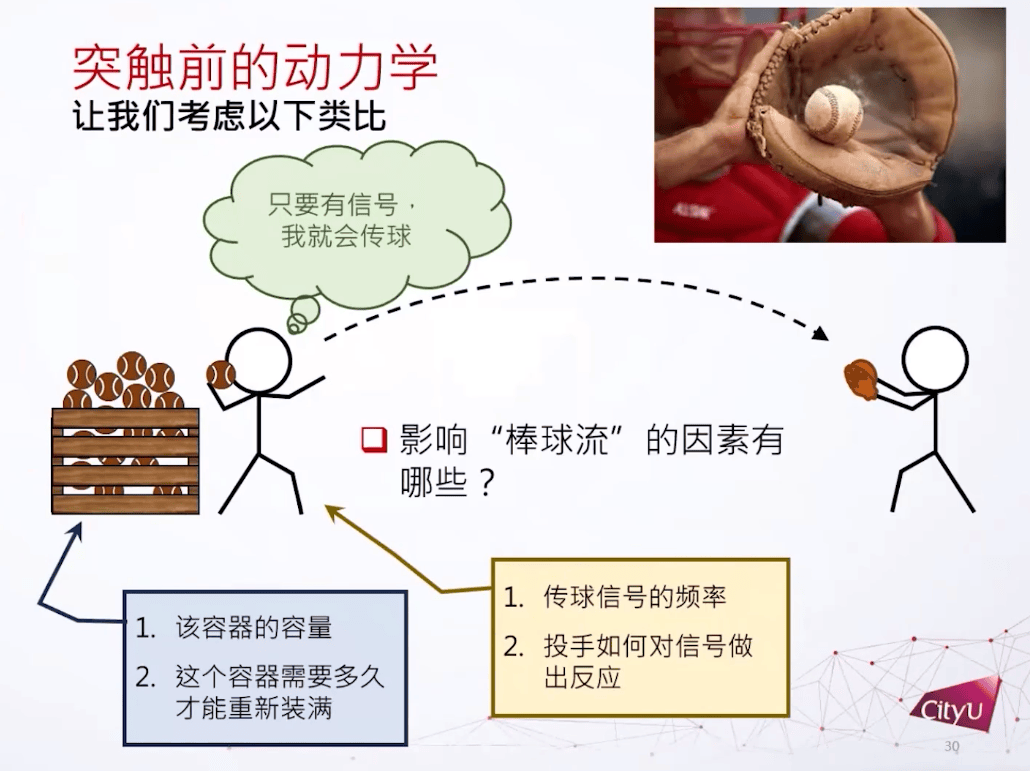
用棒球类比突触信息传递,棒球当作神经递质。这里关注的问题是,有多少库存的棒球?每次投两个还是三个?如果棒球用光了要多长时间可以重新装满?
突触动力学的实验观察
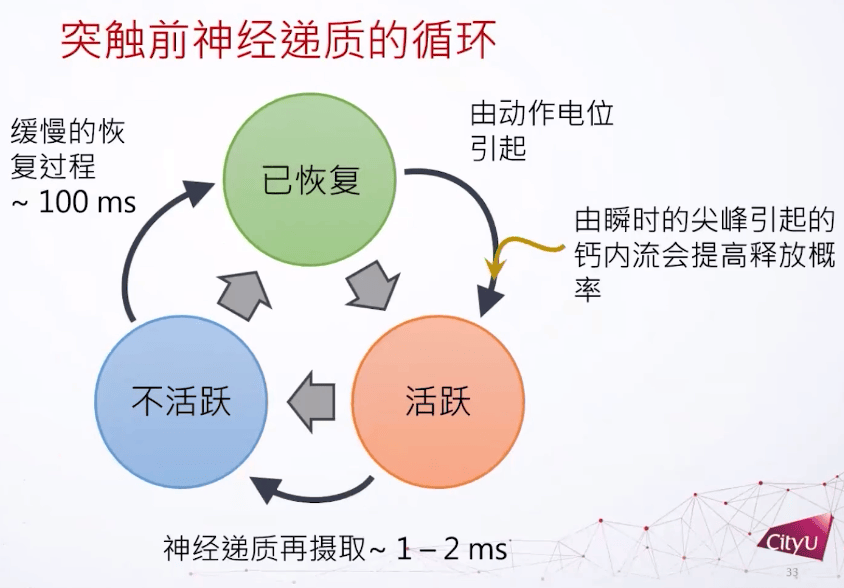
在实验中也观察到类似现象,连续的突触激发会导致突触电流的下降。神经递质发出后还有个回收的过程,神经递质不能在突触前和突触后之间存在太久,不然受体会受到过分刺激。所以只需要一瞬间打开,一两个毫秒之后就回收。神经递质定义了三个主要的态:已恢复,活跃,不活跃,它们形成一个循环。如果很久没有动作电位引起神经递质激发,就成为活跃,激发后1到2个毫秒后转为不活跃。在这个过程之后有一个比较慢的恢复过程,大概100毫秒,然后可以再激发。每次激发可以有一些钙离子的内流,可以提高释放的概率。基本上这个循环就是形容神经递质的动力学。
突触模型的描述
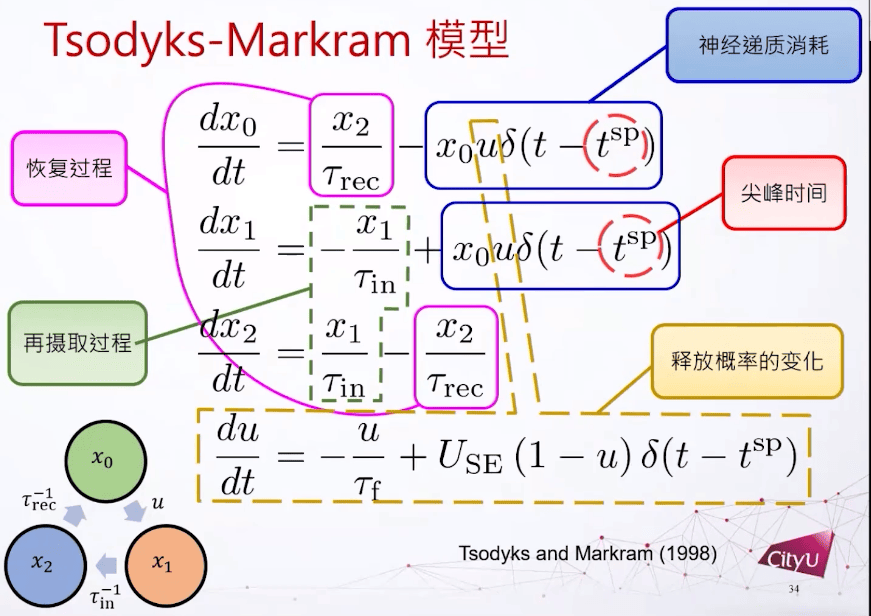
下图是突触的Tsodyks-Markram模型,下面图中框起来的6个项加起来就是0。如果我们把前三个微分方程加起来也是0,代表x0加x1加x2是一个常数。所以这里假设神经递质的总数量是一个常数。
突触的Tsodyks-Markram模型
Neural Networks with Dynamic Synapses. Misha Tsodyks, Klaus Pawelzik, Henry Markram. Neural Computation (1998).
Neural Networks with Dynamic Synapses. Misha Tsodyks, Klaus Pawelzik, Henry Markram. Neural Computation (1998).
短期突触可塑性对网络的影响
然而,短期突触可塑性对连续吸引子网络(CANN)有什么启示呢?在上世纪70年代,日本理化学研究所的甘利俊一教授已经开始提出用二元神经元(binary neuron)去模拟连续吸引子。
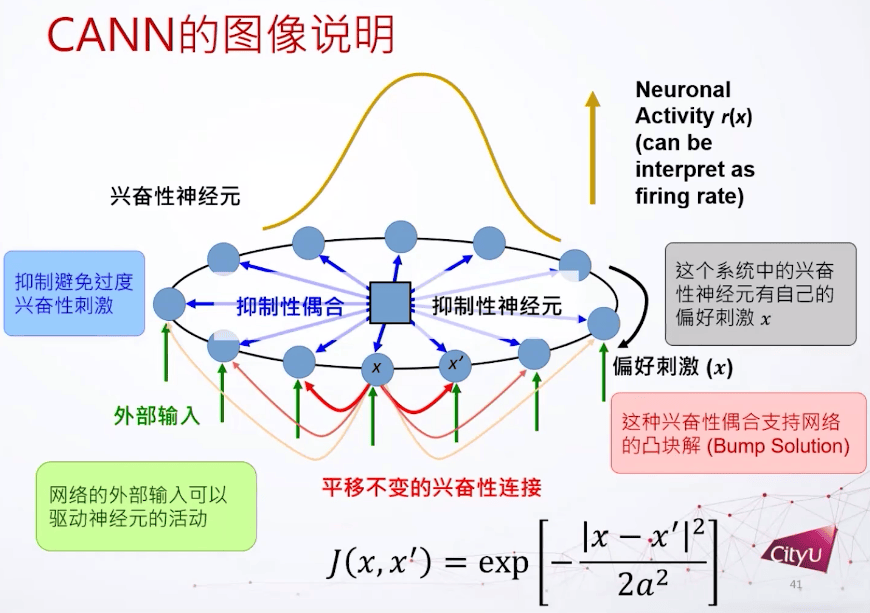
神经科学教科书里面一个比较著名的实验,猴子被训练在桌子上对不同方向做反应,做反应的运动皮层(motor cortex)要做一些编码去计划它的下一个动作。发现细胞发放率和计划的方向有关联,是一个调谐曲线,说明神经细胞有一个特定方向的偏好。
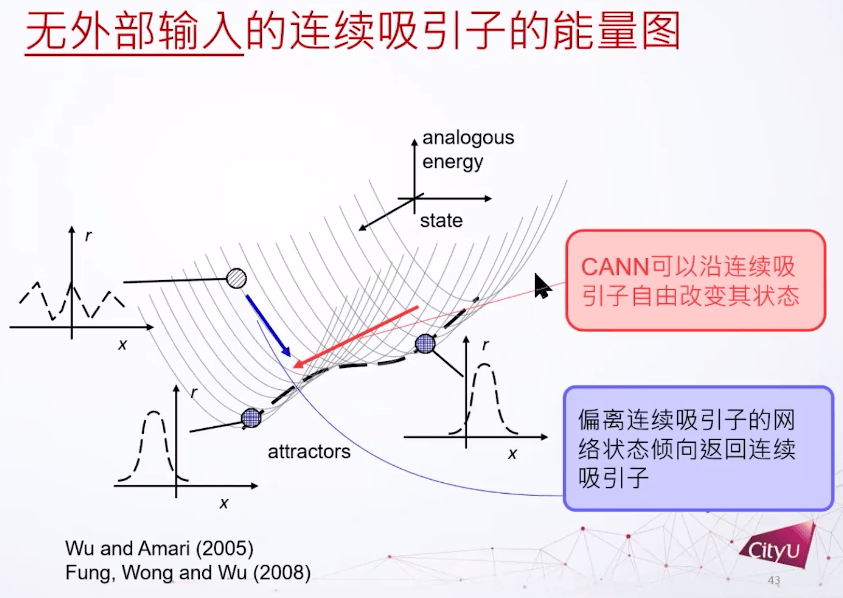
如何建模呢,假设每个细胞有自己的偏好,我们把细胞根据偏好放在环里面。兴奋性连接用高斯函数给出,还定义了抑制性的耦合和神经元,这些东西加起来可以支持一个凸块解(bump solution)。这个网络就是有了平移不变的兴奋性连接,加上一个抑制性的偶合,再加上一些外部连接,然后就可以支持这个凸块解。
我们再看连续吸引子网络的动力学。首先有个变量u,有两个参数x和t,Relaxation 项表示衰减,还有所有权重的加和,以及外部输入。
Computing with Continuous Attractors: Stability and Online Aspects. Si Wu, Shun-ichi Amari. Neural Computation
Computing with Continuous Attractors: Stability and Online Aspects. Si Wu, Shun-ichi Amari. Neural Computation
下图标出了两个稳定的态,对应的是凸块解这个解的不同位置。连续吸引子每一点都是吸引子,这个系统的态可以跑来跑去,如果偏离了这个连续吸引子的底部就会被吸回。
Dynamics of neural networks with continuous attractors. CC Alan Fung, KY Michael Wong, Si Wu. Europhysics letters.
Dynamics of neural networks with continuous attractors. CC Alan Fung, KY Michael Wong, Si Wu. Europhysics letters.
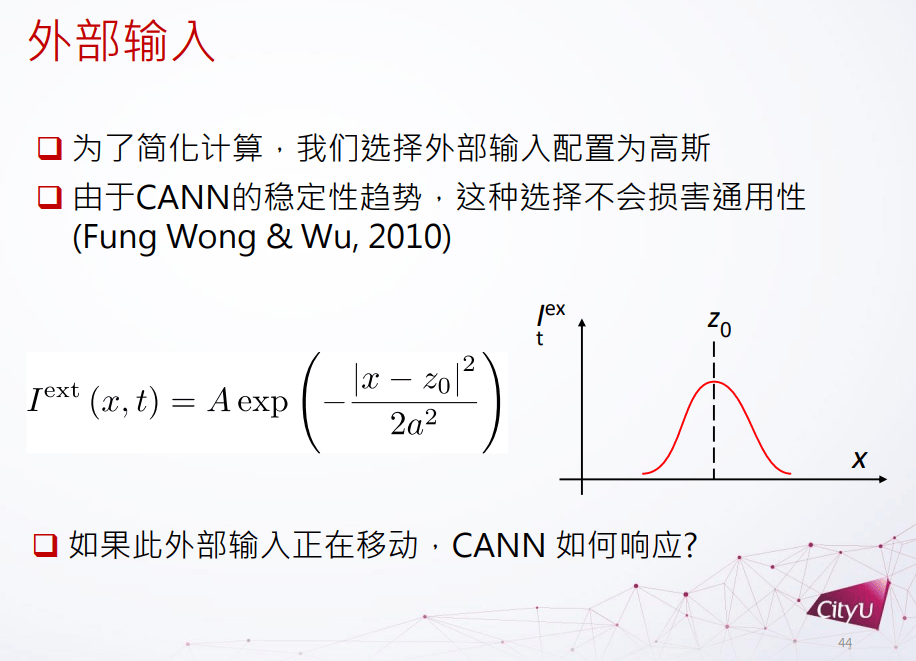
外部的输入可以有很多不同的可能性,这里简化分析外部函数为高斯函数。研究发现给高斯函数输入时连续吸引子一般不会被破坏,但是如果外部输入函数在移动,连续吸引子网络怎么反应?
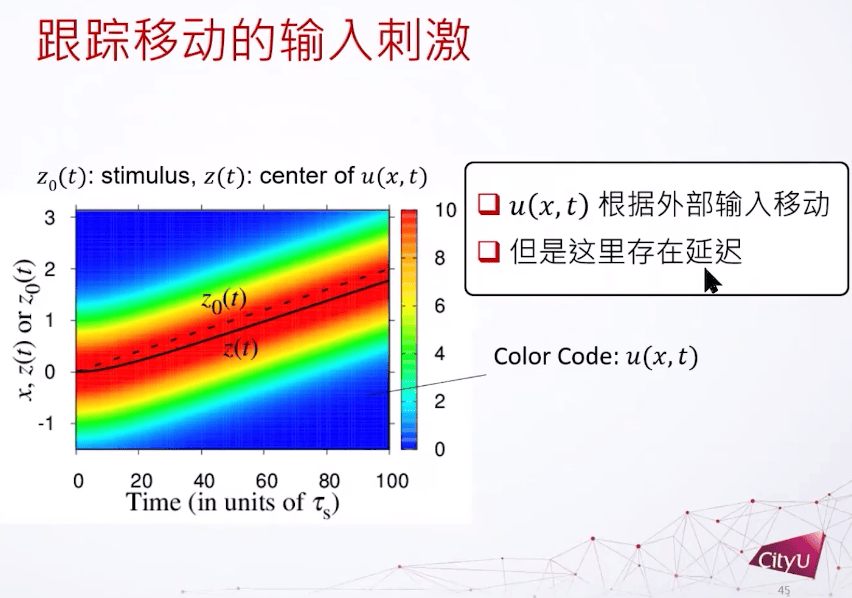
Z0是外部输入的位置,纵轴是不同输入的位置,横轴是时间。发现u会根据外部输入移动,并且有延迟。但是在大脑中,如果延迟累加以后,很可能就不能做即时的反应。
然后我们先把短期的突触动力学放进去,短期突触动力学有两个部分,一个部分是短程抑制(short term depression,STD),另外一个就是短程增强(short term facilitation,STF)。现在我们再把这个短期的衰减放进这个方程里面,本来没有p这一项,p模拟突触抑制。p这个函数会倾向于回到1这个状态,会被神经激发减小,所以结果就是如果这个神经细胞被激发,p会下降。而如果神经的活动没有减少,p会回到1。而这个τ_d这个数字它的意义就跟神经递质恢复的时间差不多,β和释放的概率差不多。这个模型和Tsodyks-Markram模型是差不多的。
Delay compensation with dynamical synapses. Chi Fung, K Wong, Si Wu. Advances in Neural Information Processing Systems.
Delay compensation with dynamical synapses. Chi Fung, K Wong, Si Wu. Advances in Neural Information Processing Systems.
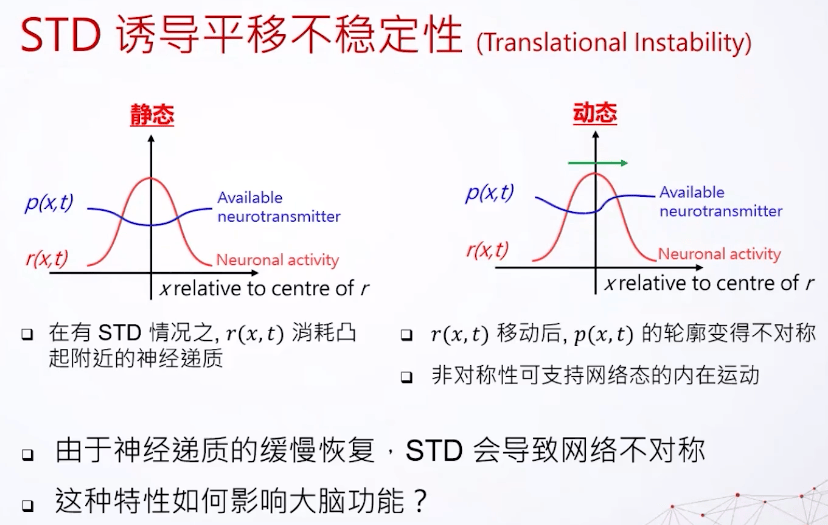
最后我们看看这个东西会怎么影响连续吸引子网络,我们只需要看它的动态跟静态。静态,比方说我们现在有一个神经的激发,他们拥有的可以激发的神经递质会减少。因为神经递质还来不及恢复,所以我们看到这里的曲线会比较小一点。动态的就不一样了,因为想象一下如果这东西在这里跑,其实是可以用到比较新的还没被完全激发的细胞,所以这里的神经递质会多一点。但是这里好像有一个尾巴,因为这是以前在上一个瞬间神经细胞活跃的区域,所以看到是变成不对称的样子。其实这是连续支持网络的动态,所以我们如果把短期的衰减加上连续吸引子网络,会有一种不稳定性,我们叫它平移不稳定性(translational instability)。是一个内在的动态,原因就是神经递质的缓慢恢复,但是这个东西怎么影响我们大脑?其实可以用来做延迟的补偿。
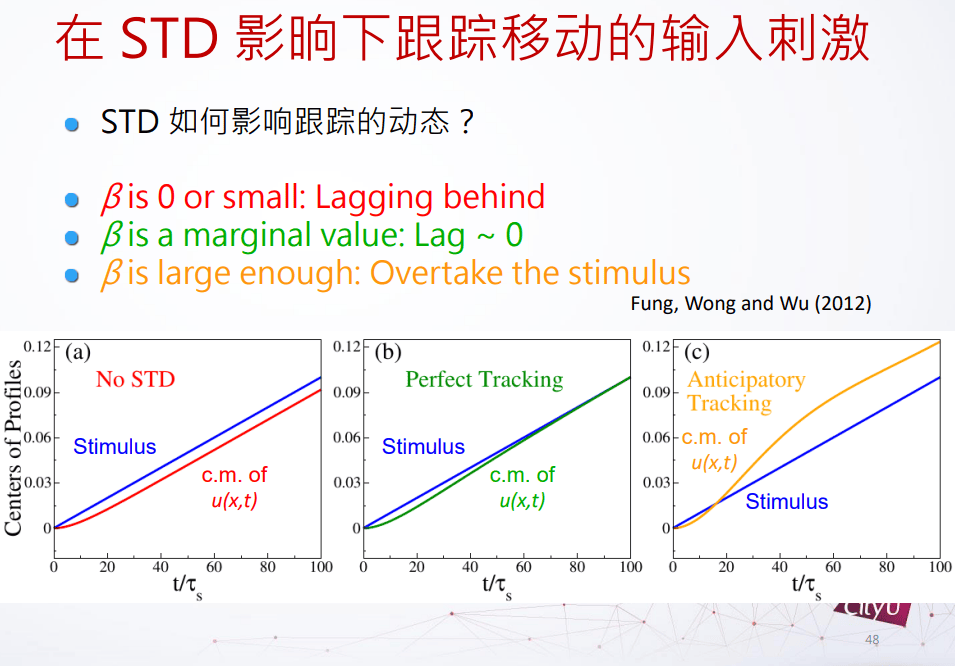
在没有短期抑制的时候,u在网络的态是落后外部刺激的,但是如果我们增加β,短期抑制力度的时候,发现延迟减少,如果再增加多一点点的时候,这个网络的态跑到移动的外部刺激的未来位置,这个东西叫它做预期跟踪(anticipatory tracking),是抵消延迟的一种可能的方案。
但是这个预期跟踪是不是真的存在?其实在我们大脑里面真的有这个。在1998年,在大鼠里面看到一些头部方向细胞指向的不是现在的头部位置,而是未来的头部位置。他们在实验当中把老鼠固定在可以转动的板子上面,之后他们转动老鼠,观察头部方向细胞的激发。现在就可以控制预期,因为得知道速率的快慢。预期性跟踪可以解释和实验上差不多的时间尺度。
Modeling Attractor Deformation in the Rodent Head-Direction System. Jeremy P. Goodridge and David S. Touretzky. Journal of Neurophysiology.
Modeling Attractor Deformation in the Rodent Head-Direction System. Jeremy P. Goodridge and David S. Touretzky. Journal of Neurophysiology.
星形胶质细胞影响短期可塑性
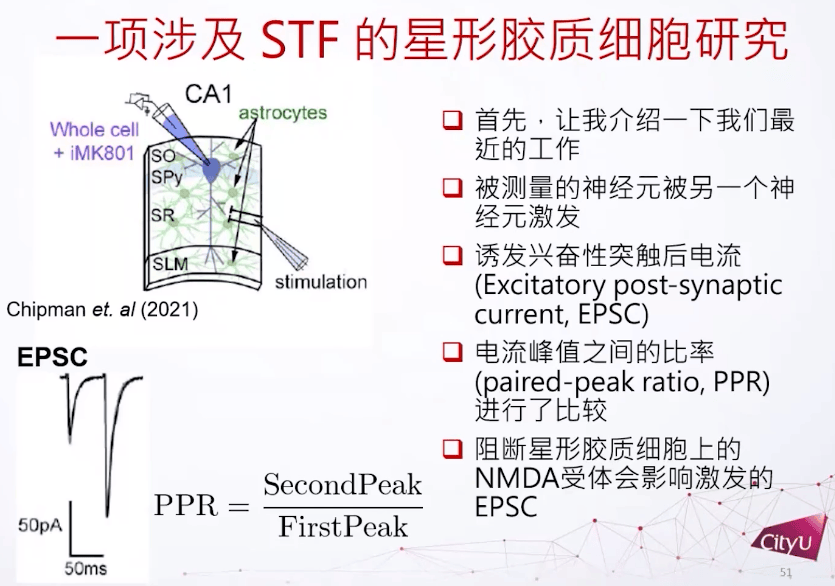
那么短期的突触可塑性和长期的突触可塑性有什么调节的关系呢?这涉及到之前的一个工作,跟星形胶质细胞有关,它是细胞旁边的胶质细胞,但是它的形状是星形的。这里有个皮层组织的切片,里面包含星形细胞和其他的神经细胞。但不是直接激发了CA1锥体细胞,而是激发另外的细胞,再间接激发CA1锥体细胞。因为想看如果影响了星形细胞的活动,这个被激发的细胞有什么影响,所以这里关注的是一个比较新的领域,就是看星形胶质细胞上面 NMDA 受体对突触后或突触前的细胞有什么影响。最主要的方法是看它的两个尖峰,因为他们每一次激发都会在突触后神经元有个尖峰。而这两个峰值的比例——PPR,是用来衡量神经元的短期可塑性有什么变化。
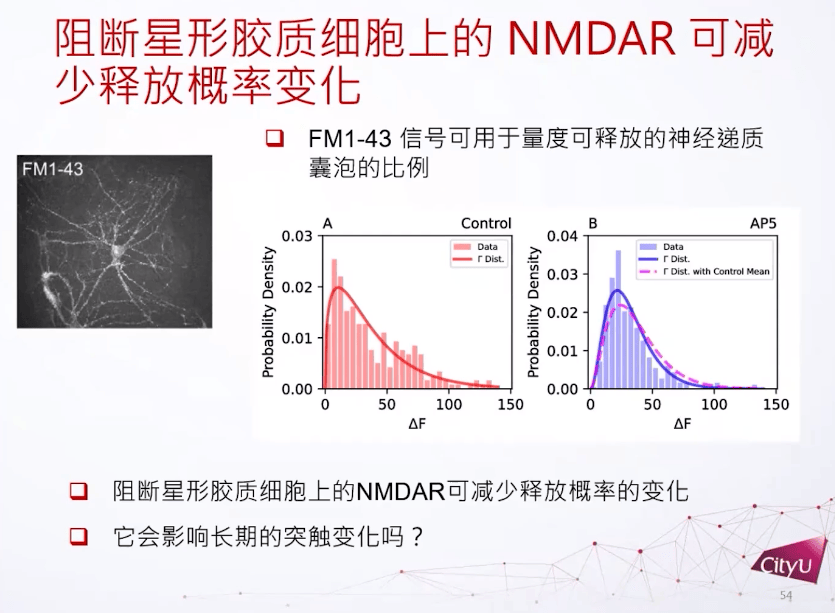
PPR是用衡量它的短期可塑性有多强的指标。比方说在这里看到第一个激发跟第二个激发其实有明显的差别,第二个激发比较强一点,所以这两个细胞是做了短期增强。如果在星形胶质细胞上再加一些NMDA受体的阻断就发现PPR比例的方差会减小,用不同的NMDA受体也是达到同样的结果。
如果我们阻断NMDA受体,在突触前其实也观察到激发概率的变化。神经递质的释放概率其实是一个伽玛分布,没有被阻断情况下,激发概率更偏向于0但是会比较长尾一点。但是如果NMDA受体被阻断,比较小的激发概率会上升,但是比较高的激发概率会减少很多。简单来说就是平均没有大的改变,但是释放概率会改变很多。
我们现在知道星形胶质细胞会影响突触后STF的变化,也会影响突触前神经递质释放概率的变化。
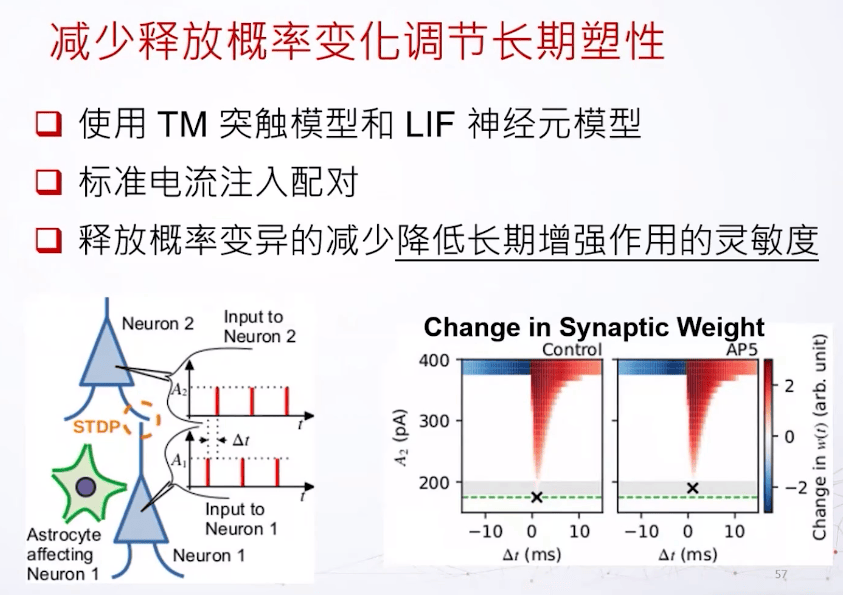
想象我们现在有两个细胞,一个突触前一个突触后。如果他的动作电位的时间有一个时间差,他们之间的耦合就会有一个变化,我们叫它突触权重,突触权重的变化是基于这个时间差到底有多大,或者是正还是负。如果突触后的发放晚于突触前,这个连接会被增强,反过来就会减弱。由于这个改变的时间持续期比较长一点,所以这个变化也称为长期可塑性。
减少释放概率的变化可以调节长期突触可塑性。可以用数值模拟做出来,我们用两个LIF(leaky integrate and fire)模型,我们先把它注入一些简单的电流。还有一个时间差,让突触前电流足够大去让它激发,但是突触后神经元输入的电流有一个不同的大小,之后我们用这个大小来判断长期可塑性的敏感灵敏度。我们知道如果突触前神经元被星形胶质细胞影响的话,它的释放概率的方差会减小,以这个观察为模型的基础,在不同的激发时间差,还有不同的突触后神经元的电流输入。可以看到如果这个星形细胞的NMDA受体被阻断,突触后神经元的敏感度会降低,因为需要更大的电流才可以激发长期突触可塑性。
使用更现实一点的泊松尖峰输入,模拟里面也是发现平均来说星形细胞的NMDA受体阻断可以调控长期突触可塑性的能力。
比较小的总结:化学突触的动力学可以重塑神经元系统的行为,首先可以破坏连续吸引子网络平移的稳定性,另外会影响长期突触可塑性。
突触竞争与模式分离
最后这个是最近的研究,突触间的竞争到底有什么用。因为在一些生物学的过程中突触其实在竞争。其中一个重要的功能就是做模式分离(pattern separation),突触的竞争提高了模式分离的能力。数据科学里经常看到不同的数据在特征空间里有重叠,在比较好的特征空间里可以把它们清晰的分开来,这是一个模式分离的过程,用简单的支持向量机就可以很好的解决。然后是成体的神经生成,这个是很大争议的问题,其实每一年都有很多人发不同文章在争论到底有没有这个事情。
比如在内嗅皮层是不是真的有神经细胞生成呢?做运动的时候,有些新细胞的反应可以观察到。不成熟的细胞在长得大一点的时候他会尝试连接不同的细胞。当成熟的时候,连接就会变得比较稳定,或者是比较有选择的连接。成熟的细胞会跟未成熟的细胞做竞争,因为成熟的细胞已经拿到连接了,新的细胞还没拿到连接,拿到稳定的位置才能继续生存下去。他们发现如果加一些药物去加强竞争,其实可以提高记忆的精度。
还没成熟细胞的突触可塑性比较强一些,在成熟之后比较弱一点。老的细胞只对一些特定的很强烈的激发有比较明显的改变。
Enhanced synaptic plasticity in newly generated granule cells of the adult hippocampus. Christoph Schmidt-Hieber, Peter Jonas & Josef Bischofberger. Nature.
Enhanced synaptic plasticity in newly generated granule cells of the adult hippocampus. Christoph Schmidt-Hieber, Peter Jonas & Josef Bischofberger. Nature.
他们还去干扰新的细胞的生成,看这个还能不能分开互相干扰的记忆。当然这个能力会消失,但是这样做其实不太会伤害神经细胞,因为神经细胞还有能力去对非常不同的记忆进行区分。但如果我们没有成体神经生成,我们区别类似事物的能力会被破坏。我们关注这个问题因为这个问题对模式分离是很重要的。
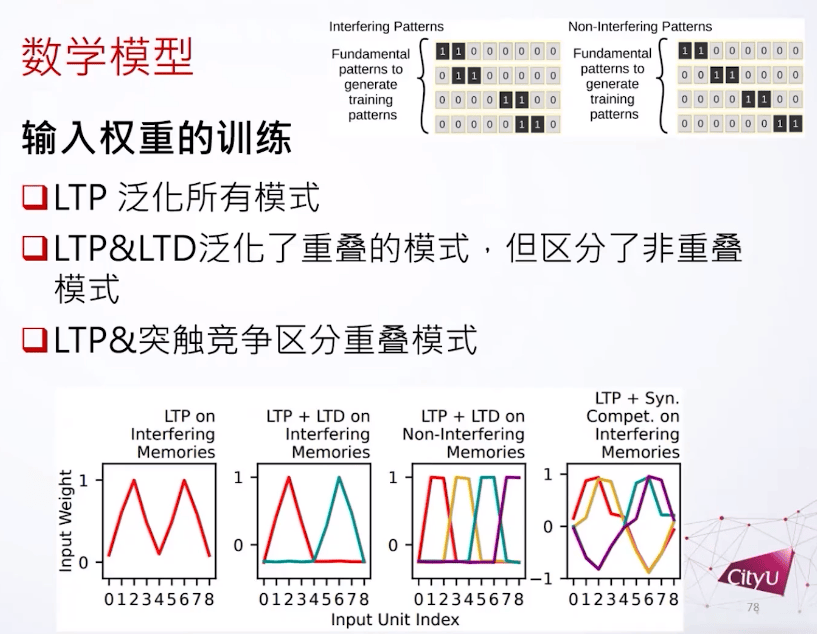
接下来做的东西有点像机器学习,首先有两种模式的数据。首先一个是干扰模式(interfering patterns),模拟一些很类似的记忆,模式之间有重叠。另外一个不干扰的模式(non interfering patterns)就是记忆之间没有关系。这个是我们用来训练的基本模式数据,再加上一个随机向量,然后生成的。他们分别训练,用来模拟不同的场景。
Competition on presynaptic resources enhances the discrimination of interfering memories. Chi Chung Alan Fung, Tomoki Fukai. PNAS Nexus 2023.
Competition on presynaptic resources enhances the discrimination of interfering memories. Chi Chung Alan Fung, Tomoki Fukai. PNAS Nexus 2023.
输入单元是0-10的数字。我们现在要学的就是到底什么样的权重帮可以模式识别。
在这里为什么突触竞争重要呢,我们看如下三个场景:
我们现在是模拟的不是尖峰神经元,用的是基于发放频率的神经元,所以我们用赫布学习来模拟LTP。
如果我们用LTP来训练,我们发现神经细胞会对所有的模式都有反应,我们说它泛化了所有的模式。如果LTP加LTD,看到神经细胞分成两组,但是它还是不能分别重叠的模式。但是LTP加上突触竞争,很奇怪的是他们可以区别所有模式了。
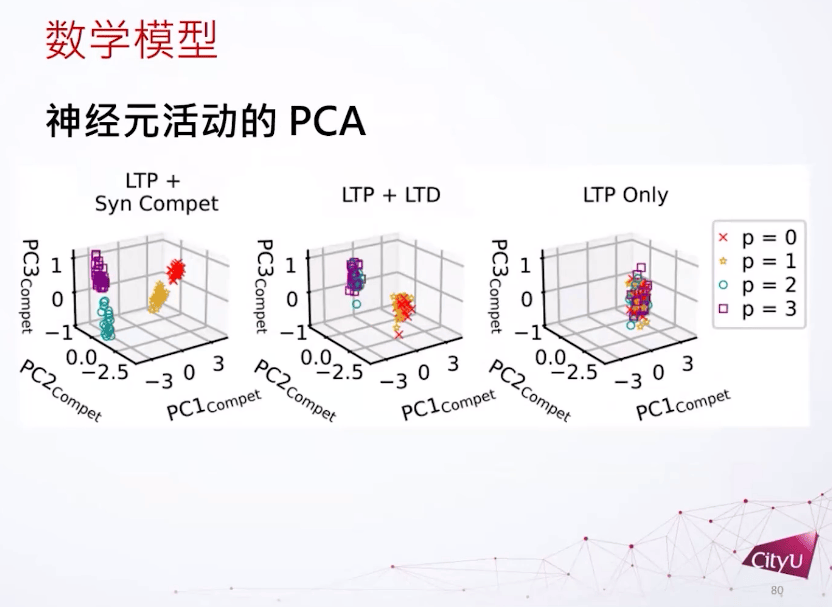
把这个输入权重用PCA分类。如果我们只看这个LTP,所有特征都会重叠,因为倾向于把所有东西都泛化,但是LTP加上突触竞争可以分离所有模式。
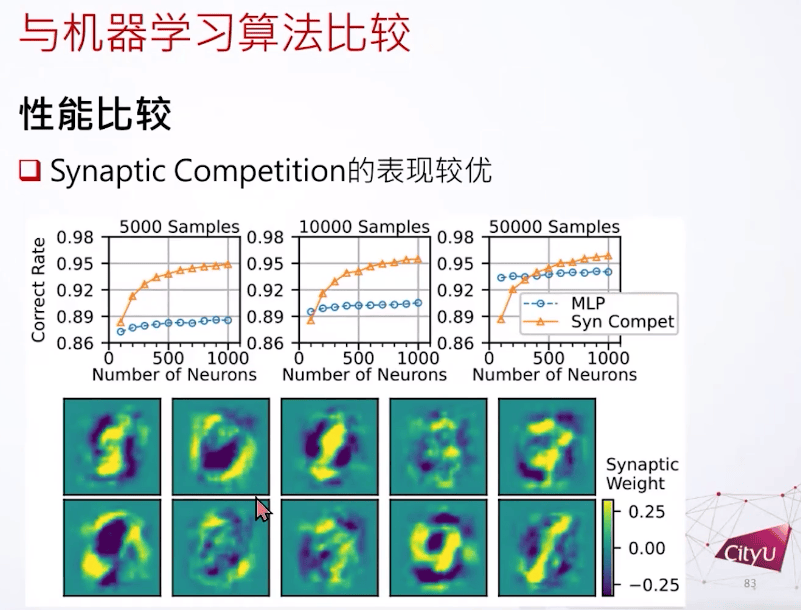
之后我们在MNIST数据集里测试。我们发现如果用很少的样例,还有短一些的训练时间,突触竞争是可以做到比较好的结果。比如突触竞争只学了一次,但结果是让人意外的好。学一次就能泛化出这些很奇怪的输入权重,好像是把溶解的数字放进他们的输入权重,或者是自动把这个突触竞争搞成一个特征,把不同特征放在不同的神经元一样。
这个是第二个总结就是这是基于竞争的学习规则,来自成体神经生成,但在我们的不同场景。比较少资源的时候其实这个这个突触竞争比反向传播要好一点。很奇怪的是这个输入的权重会看起来像溶解的数字一样,好像把不同特征分叉开来。在这个应用上我们看到突触竞争可以支持模式分离。
|学者简介
冯志聪(Alan Fung),2006 年开始研究连续吸引子神经网络(CANN)模型。
研究方向:专注在计算神经科学领域,基于数学工具和数值方法的基础脑科学的计算模型、神经现象的机器学习实现、数据驱动的神经数据分析,希望能够促进神经科学和人工智能的相互启发。
本文来源:集智俱乐部











内容来自网友分享,若违规或者侵犯您的权益,请联系我们
所有跟帖: ( 主贴楼主有权删除不文明回复,拉黑不受欢迎的用户 )
进入内容页点击屏幕右上分享按钮
楼主前期社区热帖:
- 广东进入登革热流行季,省疾控局做出三点提醒 09/22/24
- 妊娠期易被忽视的补剂!缺乏可增加先心病、自闭症等风险 09/22/24
- 求知探索,格物穷理 09/22/24
- 谋大事者,首重格局 09/22/24
- 苏轼与陇馔之“熊腊” 09/22/24
- 王阳明10句话,心学精华,人生要旨 09/20/24
- 又是人间一度秋,层林尽染万山红 09/20/24
- 这些成语都出自古建 09/20/24
- 这些成语都出自古建 09/20/24
- 有酒有味,自在生活,四川人把“松弛感”玩明白了 09/20/24
- 四川人说的“骚鸡”不是拿来吃的,却伴随我们上千年,智慧的结晶 09/20/24
- 小璐医生话代谢 | 秋分将至,老年人如何用药食同源的麦冬治失眠 09/20/24
- 【世界阿尔茨海默病日】警惕!出现这10个症状,可能是痴呆的早期信号! 09/20/24
- 【全国爱牙日】从摇篮到摇椅,如何守护牙齿健康? 09/20/24
- 提醒:3类运动是为膝盖“上刑”,要避免!医生分享五个护膝方法 09/18/24
- 关于淋巴瘤的这些误区,您知道多少? 09/18/24
- 泸州有一种味道,不是非遗胜似非遗,吃过的人才懂得什么是幸福 09/18/24
- 古代玫瑰的那些故事 09/18/24
- 读懂《菜根谭》经典十句,人生豁然开朗 09/18/24
- 长知识丨“客气”原来是这样来的 09/18/24
>>>>查看更多楼主社区动态...