
连系新冠病毒与Ralph S. Baric的双纽带(下)(续)
| BY: 苦难与荣耀 |
接续:连系新冠病毒与Ralph S. Baric的双纽带(下)
https://web.6parkbbs.com/index.php?app=forum&act=view&tid=4253444
6pa论文B-4 6park.com2008年9月1日(Published online 2008 Jun 25),Ralph S. Baric等人在Journal of Virology发表了论文:Pathways of Cross-Species Transmission of Synthetically Reconstructed Zoonotic Severe Acute Respiratory Syndrome Coronavirus(人畜共患严重急性呼吸综合征冠状病毒的跨物种传播途径)
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2519660/
https://journals.asm.org/doi/10.1128/JVI.00818-08 6park.comRalph S. Baric依然是论文的通讯作者。 6park.com

病毒1:嵌合病毒icSZ16-S,它的刺突蛋白来自果子狸冠状病毒SZ16,骨架来自SARS病毒株Urbani,
病毒2:icSZ16-S K479N,它是对icSZ16-S的479氨基酸实施人为替换(K479N)后得到的病毒,
病毒3:icSZ16-S K479N D22,它是icSZ16-S K479N在人气道上皮上培养、传代、筛选得到的实验室变异体,
病毒4-6:三个SARS病毒的克隆毒株:icSARS、icCUHK-W1、icGZ02,
病毒7:SARS不同毒株的人工混嵌体icGD03-S,它应该是SARS-GD03的刺突蛋白与SARS-Urbani的骨架嵌合而成的。 6park.com关于病毒4-6的更多说明:
icSARS是用反向遗传平台合成的SARS-Urbani的克隆,SARS-Urbani是SARS疫情晚期分离自美国的SARS病毒株。
GZ02是SARS疫情早期分离自广州的SARS病毒株。
CUHK-W1是SARS疫情中期分离自香港的SARS病毒株。
GD03是比Urbani更晚的SARS病毒株,它是2004年1月疫情再次零星出现时分离自一散发广东患者的SARS病毒株。
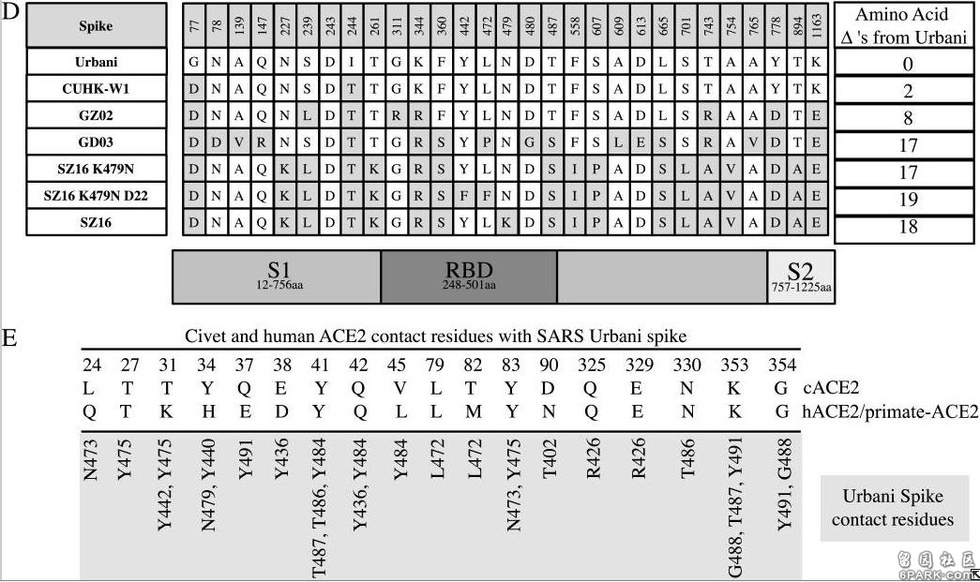
(SARS疫情发生于2002年11月16日-2003年9月2日,高峰期为2002年11月16日-2003年7月16日) 6park.com论文详尽地列出了所涉及各病毒刺突蛋白的氨基酸差异,并标出了其它6种刺突蛋白相对于SARS-Urbani刺突蛋白的氨基酸差异数,见图D;
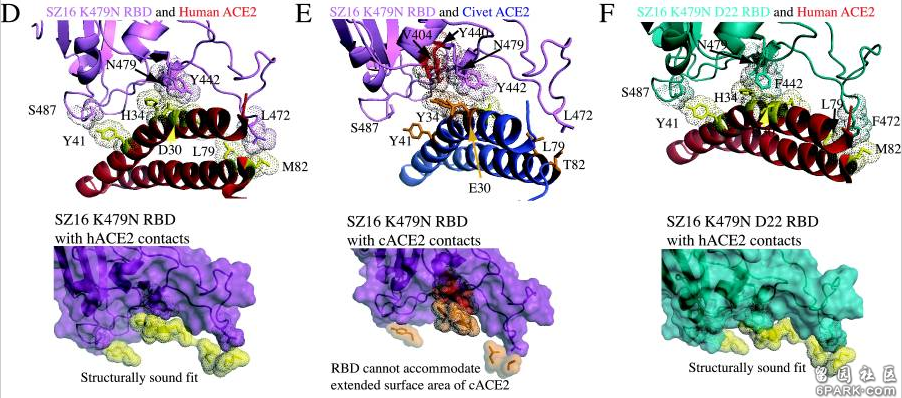
论文还细致地列出了cACE2(civet ACE2)及hACE2或primate-ACE2(灵长类动物ACE2)的所有热点氨基酸残基及与它们发生作用或接触的SARS-Urbani的RBD氨基酸残基,见图E。 6park.com
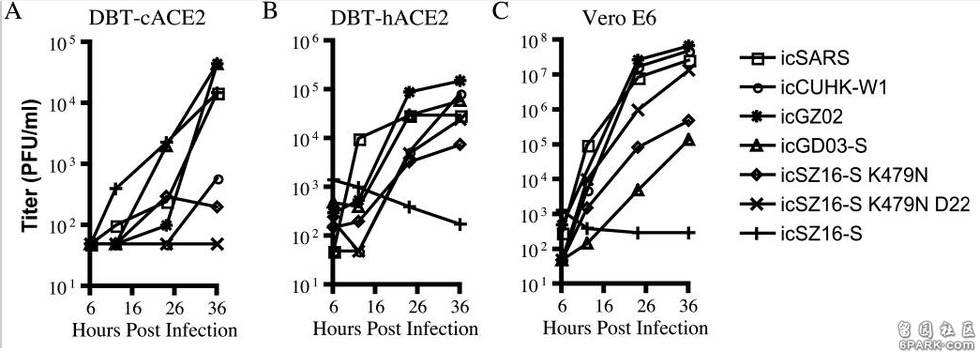
7种病毒对4种细胞中的感染、复制实验数据图。
横坐标是感染后时段,单位为小时;纵坐标是病毒的复制滴度,单位为PFU/ml。
以上实验数据图表明:
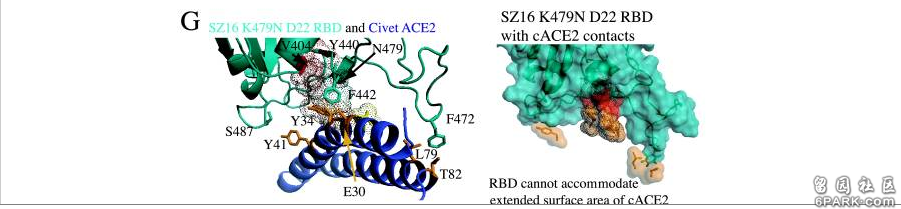
a)icSZ16-S K479N、icSZ16-S K479N D22及icCUHK-W1不能在DBT-cACE2细胞中有效复制,或只能低水平复制,而其它病毒,包括icSZ16-S都能在DBT-cACE2细胞中有效复制,见图A; 6park.comb)icSZ16-S不能在DBT-hACE2细胞中有效复制,而其它病毒都能在DBT-hACE2细胞中不同程度地复制(K479N复制能力相对较差),见图B; 6park.comc)icSZ16-S不能在Vero E6细胞中有效复制,而其它病毒都能在Vero E6细胞中不同程度地复制,GZ02、CUHK-W1、icSARS、icSZ16-S K479N D22在Vero E6细胞中的复制都极为强烈,见图C; 6park.com(论文还指出:上述七种病毒皆不能感染未转基因的普通小鼠DBT细胞。) 6park.com可见,icSZ16-S与icSZ16-S K479N及icSZ16-S K479N D22在细胞感染、复制能力,物种ACE2结合能力上存在巨大差异。 6park.comicSZ16-S、icSZ16-S K479N二者的刺突蛋白只有一个氨基酸不同,即第479氨基酸,icSZ16-S的第479氨基酸是赖氨酸K,而icSZ16-S K479N的第479氨基酸被人为替换为SARS-Urbani的第479氨基酸--天冬酰胺N(人为制造了K479N突变)。刺突蛋白第479氨基酸是第三个RBD关键氨基酸。 6park.comicSZ16-S、icSZ16-S K479N D22二者的刺突蛋白有三个氨基酸不同,除了K479N差异外,它们还有另外两个氨基酸差异:Y442F和L472F。刺突蛋白第442、472氨基酸分别是第一、第二个RBD关键氨基酸。 6park.com
相关病毒RBD关键氨基酸对照表
icSZ16-S、K479N、K479N D22这三个病毒在不同物种ACE2结合能力,不同物种细胞感染、复制能力上的巨大差异就是它们RBD关键氨基酸的差异造成的。 6park.com为了解释各病毒与cACE2、hACE2的不同结合能力,研究者使用RosettaDesign和Modeler软件开发了4种病毒(SARS-Urbani、SZ16、icSZ16-K479N、SZ16-K479N D22)刺突蛋白与cACE2、hACE2相互作用的结构模型,深入分析了各病毒与cACE2、hACE2在各个关键氨基酸位点的作用情况,解读了关键氨基酸差异对hACE2结合能力的影响。论文指出: 6park.com1)cACE2分子与hACE2分子有两个关键残基(E30和Y34)不同,cACE2的E30和Y34的附加甲基可能会使cACE2结合界面产生明显的突起。SARS-Urbani的RBD能够良好适应hACE2的结合界面,见图A;同时,它又能容纳cACE2的表面突起,见图B。因此,SARS-Urbani对人类和civet(果子狸或麝香猫)有双重宿主适应能力; 6park.com
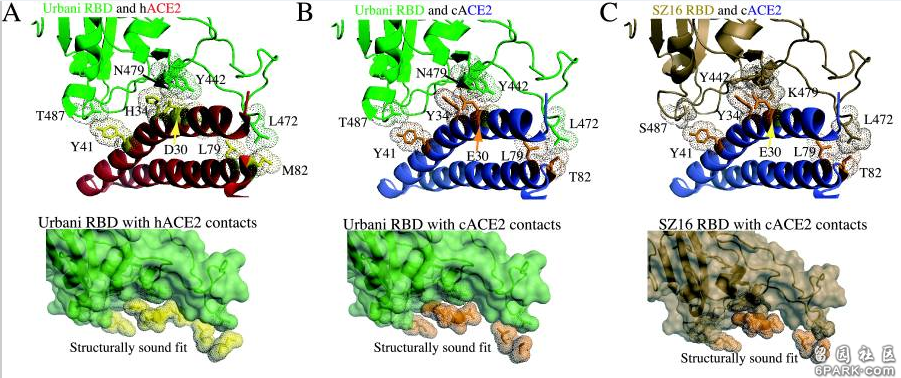
2)与Urbani RBD类似,SZ16 RBD也能容纳cACE2的E30、Y34造成的表面突起,以实现有效结合,见图C; 6park.com3)SZ16 K479N中的K479N突变(人为替换)引入的479N(天冬酰胺)重塑了SZ16刺突蛋白的结合界面,促进了其与hACE2的结合,见图D; 6park.com
早在2007、2008年,Ralph S. Baric对冠状病毒的RBD关键氨基酸,对它们与ACE2,特别是与hACE2的作用机制,就已经研究得极为深入、透彻了;
早在2007,Ralph S. Baric就已经成功实施了关键氨基酸的人为替换,成功实施了冠状病毒的人类组织适应性培养,成功改造、培养出了能感染人体细胞的实验室病毒。 6park.com本系列三篇文章结论
1)有人参照WIV1(rs3367)病毒的关键氨基酸,通过氨基酸复用及等价、等效替换,为新冠病毒设计了一套与WIV1的关键氨基酸高度等价、等效的关键氨基酸。 6park.com2)新冠拥有与WIV1高度等价、等效的关键氨基酸,导致了以下“巧合”:
a)新冠、WIV1的跨物种感染能力都极为强大,难分伯仲;
b)新冠、WIV1的宿主范围都极为广泛,而且,它们的宿主范围高度雷同(它们可感染的物种高度重合);
c)新冠、WIV1的刺突蛋白都极为适宜结合hACE2,与hACE2的结合亲和力都极为强大。 6park.com3)新冠病毒关键氨基酸的设计者,即新冠病毒的设计者,极有可能就是Ralph S. Baric,或者Ralph S. Baric团队的某个、某些资深成员。
贴主:hgao于2024_03_26 13:32:12编辑
喜欢hgao朋友的这个贴子的话, 请点这里投票,“赞”助支持!
请点这里投票,“赞”助支持!

内容来自网友分享,若违规或者侵犯您的权益,请联系我们
所有跟帖: ( 主贴楼主有权删除不文明回复,拉黑不受欢迎的用户 )
hgao兄,看到了你的来信,请你不要再转发我的文章了。 - 苦难与荣耀 (27 bytes) 03/26/24
.... - hgao (46 bytes) 03/26/24
请hgao兄在文章中注明原文创作时间,并提供原文地址。 (无内容) - 苦难与荣耀 (0 bytes) 03/26/24
桂花酒 给 hgao 点“赞”支持3银元奖励!! (无内容) - 桂花酒 (0 bytes) 03/26/24
hgao 给 桂花酒 点“赞”支持3银元奖励!! (无内容) - hgao (0 bytes) 03/26/24
打开微信,扫一扫[Scan QR Code]
进入内容页点击屏幕右上分享按钮
进入内容页点击屏幕右上分享按钮
楼主前期社区热帖:
- 顶不住啊顶不住!蓝燕,这身材谁能顶得住嘛 04/26/24
- 原来气血才是女人的命根子!气血足、卵巢好对女人竟这么重要… 04/26/24
- 梁从诫回忆母亲《我的母亲林徽因》重新认识这位民国著名才女 04/26/24
- 多走路的人能長壽?北大最長壽的教授,一語點破養生精髓:你以為是在 04/26/24
- 服用中药之后,需要多久时间才见效?医生:一般有3个级别 04/26/24
- 澳洲移民故事:在澳洲金矿工作的武汉小伙,年薪你大胆猜一猜 04/26/24
- 听劝, 受不了这7件事, 千万别移民澳洲! 来了肯定后悔! 澳洲已经入睡了 04/26/24
- 海外友人回国的适应和不适应,春节断亲成潮流,说明了什么 04/26/24
- 6年没回国的我终于回去了!国内国外的差异越来越大 04/26/24
- 提醒:絕不能放進微波爐的10樣東西,很多人還不知道 04/26/24
- 癌症逆轉怎麼做到?哈佛醫師教你5招防癌:10年不復發 04/26/24
- 催眠音乐432Hz- Alpha Waves Heal The Whole Body and ... 04/26/24
- 血糖實測報告 那些看似健康的可怕食物 ?這些吃多注定糖尿病? 04/26/24
- 比糖更危險的一種碳水,吃了很久卻不知道 04/26/24
- 十種破壞腎臟食物排行榜,不知不覺腎功能下降的比同年齡的更快 04/26/24
- 腎臟老太快?慢性腎衰竭?十大常吃護腎食物搶救你的腎功能 04/26/24
- 外蒙古是如何被丟掉的,誰是千古罪人? 一口氣看完蒙古獨立全過程 04/26/24
- 育空淘金第二季2:淘金小白第一年淘金就挖到14公斤重的黄金,二代 04/26/24
- 育空淘金客S1:四队淘金小白怀揣着淘金梦来到育空淘金,看他们谁能发大财谁赔 04/26/24
- 警惕男人婚前承诺!年轻夫妻婚前老公答应生完小孩不用带 婚后却大变样 04/26/24
>>>>查看更多楼主社区动态...